

アミノ酸と尿素回路
代謝全体マップは以下図参照。

アミノ酸とは
アミノ酸とはアミノ基とカルボキシル基を持つ化合物であり、カルボキシル基に結合する炭素をα炭素と呼び、α炭素にアミノ基が結合したものをαアミノ酸と呼ぶ。
生体内には20種類のアミノ酸があるが、全てがαアミノ酸で構成されている。
これらのαアミノ酸のうちグリシン以外は光学異性体を持ち、L体のみがタンパク質合成に利用される。タンパク質は多数のアミノ酸がペプチド結合(アミド結合)によりつながったものです。
人の体の中ではリボソームでmRNAの情報を読み取り、体の中のアミノ酸を使いタンパク質が合成されることは細胞の構造の項でお話したとおりですが、この時タンパク質合成に使われるアミノ酸は食物として外部から摂取しなければなりません。
とは言ってもすべてのアミノ酸を外部から摂取しなければならないわけではありません。たとえばシステインはメチオニンから作られるし、チロシンはフェニルアラニンから作られます。
このようにアミノ酸の中でも元となるアミノ酸のみを摂取する必要があります。
必須アミノ酸
この元となるアミノ酸の中でも、体内で合成できないアミノ酸のことを必須アミノ酸と呼びます。(語呂:トロリバスフメイorふろばいすひとりじめ)
- 必須アミノ酸・・・ロイシン、イソロイシン、バリン、リシン、トレオニン、トリプトファン、メチオニン、フェニルアラニン
- 非必須アミノ酸・・・ヒスチジン、アルギニン、アラニン、システイン、チロシン、プロリン、セリン、グリシン、アスパラギン酸、アスパラギン、グルタミン酸、グルタミン
分岐鎖アミノ酸(BCAA)
バリン、ロイシン、イソロイシンは分岐鎖アミノ酸(Branched Chain Amino Acid)と呼ばれ、側鎖が枝分かれしているのが特徴である。
BCAAは必須アミノ酸の約40%を占め、筋肉のエネルギー源として、特にロイシンは顕著に筋タンパク分解を抑制し、タンパク合成を促進する。
肝不全時にはBCAAが低下して、肝臓で代謝されるAAA(芳香族アミノ酸)が上昇することによりフィッシャー比(=BCAA/AAA)が低下する。(リーバクトの適応)
糖原生アミノ酸とケト原生アミノ酸
純粋なケト原生アミノ酸はロイシンとリシンのみで、糖原生とケト原生の両方の性質を持っているのがイソロイシン、フェニルアラニン、チロシン、トリプトファン、トレオニン、それ以外が純粋な糖原生アミノ酸です。
糖原生アミノ酸は飢餓時等において、糖新生(肝・腎で)によりグルコースへ戻れるアミノ酸で、ケト原生アミノ酸は代謝されてアセチルCoAもしくはケトン体となった後アセチルCoAに戻されて、エネルギー産生に回るアミノ酸です。
糖代謝のところでも述べた通り、ケト原生アミノ酸がアセチルCoA経由でTCA回路に入って糖新生に回る(糖原生はない)ことはないので注意する。理由は糖新生に必要なオキサロ酢酸をTCA回路に入るのに使えないためです。
各アミノ酸の作用
- アラニン(Ala)はアミノ基転移反応でαケト酸であるピルビン酸に変換される。
- グルタミン(Gln)はグルタミナーゼの作用でアンモニアを遊離し、グルタミン酸となり、アミノ基転移反応でグルタミン酸デヒドロゲナーゼとNAD+の作用でα-ケトグルタル酸へと変換される。代わりにアスパラギン酸とアラニンが生成する。末梢のアンモニアを肝臓に運んで除去するルートとして、グルタミンシンテターゼにより遊離のアンモニアを結合し、グルタミンとして2つのアミノ基を運ぶルートが重要。
- ヒスチジン(His)はアンモニアを遊離してグルタミン酸(Glu)に変換される。ヒスチジンからはVB6+ヒスチジンデカルボキシラーゼの作用でヒスタミンが、グルタミン酸からはVB6+グルタミン酸デカルボキシラーゼの作用でガンマアミノ酪酸(GABA)が生成される。
- アルギニン(Arg)は加水分解してオルニチンを生成後、グルタミン酸5-セミアルデヒドを経て、グルタミン酸(Glu)に変換される。アルギニンは一酸化窒素合成酵素によりNADPH存在下NOを遊離する(神経、マクロファージ、血管内)
- プロリン(Pro)はグルタミン酸5-セミアルデヒドを経て、アルギニンと同じルートでグルタミン酸に変換される。
- トレオニン(Thr)はグリシン(Gly)に変換され、グリシンはセリン(Ser)に変換されて、アンモニアを遊離し、ピルビン酸に変換される。また、トレオニンは別経路でアセチルCoAを遊離してグリシンに変換される。
- システイン(Cys)は複数の経路でピルビン酸に変換される。
- トリプトファン(Trp)はキヌレニン経路でアセト酢酸を生成する間に、アラニンを遊離するのでピルビン酸にも変換される。トリプトファンからはセロトニンが生成される。
- アスパラギン(Asp)はアスパラキナーゼの作用でアンモニアを遊離し、アスパラギン酸(Asp)となり、アミノ基転移反応でαケト酸であるオキサロ酢酸に変換される。
- イソロイシン(IIe)とバリン(Val)、メチオニン(Met)はプロピオニルCoA経由でスクニシルCoAに変換後TCA回路に入り糖新生などに利用される。イソロイシンがプロピオニルCoAに変換される際にアセチルCoAが生じる。
- ロイシン(Leu)はアセチルCoAとアセト酢酸を生じる。
- メチオニンはS-アデノシルメチオニン(SAM)に変換され、SAMはグアニジノ酢酸と反応してクレアチニンを合成する。またSAMはホモシステインとなって、VB12と5-メチルテトラヒドロ葉酸と反応し、メチオニンに再生されるか、セリンと縮合してシスタチオン経由でシステインの合成及びαケト酪酸からのTCA回路へ入る。またSAMはノルアドレナリンにメチル基を与えてアドレナリンにする際にも使われる。
- リシン(Lys)はサッカロビン経由でアセト酢酸へと合成され、アセチルCoAに戻される。
- フェニルアラニン(Phe)はチロシン(Tyr)経由でフマル酸とアセト酢酸に変換される。フマル酸はそのまま、アセト酢酸はアセチルCoAに戻りTCA回路か脂肪酸合成へ。フェニルアラニンとチロシンはドパミンやノルアドレナリンを生成する。
尿素回路(オルニチン回路)
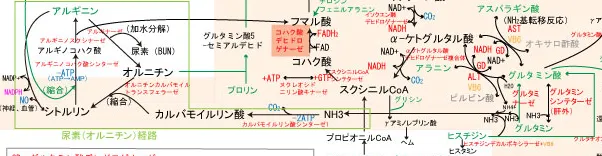
尿素回路(オルニチン回路)は肝臓のミトコンドリアと細胞質にまたがる、アミノ酸のアミノ基がアンモニアに変えられた後に、尿素に変換して排泄するための回路である。
末梢組織含めアミノ酸のアミノ基のほとんどはアミノ基転移反応でグルタミン酸に集められ、グルタミン酸がαケトグルタル酸に変換される脱アミノ反応でアンモニアとして放出される。
末梢組織には尿素回路がないため、末梢組織で生じたアンモニアはグルタミンサンシンテターゼによってグルタミン酸に結合し、グルタミンとなって血液に入り、肝臓まで移動し、肝臓でグルタミナーゼの作用でグルタミン酸に戻される過程でアンモニアとして放出される。
これらのアンモニアは肝臓のミトコンドリア内でカルバモイルリン酸シンターゼの活性によって二酸化炭素と結合し、カルバモイルリン酸を形成する。この反応が尿素回路の律速であり、2ATPを使用する。
カルバモイルリン酸は、オルニチンカルバモイルトランスフェラーゼの活性によりオルニチンと縮合し、シトルリンを生成する。
カルバモイルリン酸シンターゼとオルニチンカルバモイルトランスフェラーゼの2つの酵素はミトコンドリアマトリックス内に存在している。
生成されたシトルリンは細胞質へ移動し、アルギノコハク酸シンターゼの活性によりアスパラギン酸と縮合し、アルギノコハク酸が生成する。この際ATPが使用されるが、ATP→ADPではなく、ATP→AMPでありATPが失うリン酸基が2つである点が重要。
アルギノコハク酸はアルギノスクシナーゼの活性によりアルギニンとフマル酸に別れ、フマル酸はTCA回路へ、アルギニンはアルギナーゼの活性により加水分解されてオルニチンと尿素(BUN:血中尿素窒素ともいう)を生成する。
尿素の2つのアミノ基の1つはアンモニア由来、もう一つはアスパラギン酸由来である。
なお、アルギニンはグリシンと反応してグアニジノ酢酸(クレアチン生成の中間体)やオルニチンを生成する反応にもかかわる。クレアチンは筋肉に大量に存在するクレアチンキナーゼ(CK)とATPを使ってクレアチンリン酸の形で蓄えられている。
全体の反応は、NH3+H2O+C02+アスパラギン酸+3ATP → 尿酸+フマル酸+2ADP+2Pi+AMP+PPi。
参考サイト:薬学部学生向け講義資料ダウンロード
- 糖
- 脂質
- タンパク質
-
ビタミン
├ 脂溶性ビタミン
| ├ ビタミンA
| ├ ビタミンD
| ├ ビタミンE
| ├ ビタミンF
| └ ビタミンK
|
└ 水溶性ビタミン
├ ビタミンB1
├ ビタミンB2
├ ビタミンB6
├ ビタミンB12
├ ビタミンC
├ ビタミンM
├ ビタミンH
├ ビタミンP
├ ビタミンQ
├ ニコチン酸
└ パントテン酸
コメントor補足情報orご指摘あればをお願いします。
- << 前のページ
- 次のページ >>
